
Сигнал распознавания частицы РНК - Signal recognition particle RNA
| RN7SL1 | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||||||||||||||||
| Псевдонимы | RN7SL1 , 7L1a , 7SL, RN7SL, RNSRP1, РНК частицы распознавания сигнала, РНК, 7SL, цитоплазматическая 1, РНК-компонент частицы распознавания сигнала 7SL1 | ||||||||||||||||||||||||
| Внешние идентификаторы | OMIM : 612177 GeneCards : RN7SL1 | ||||||||||||||||||||||||
| |||||||||||||||||||||||||
| Ортологи | |||||||||||||||||||||||||
| Разновидность | Человек | Мышь | |||||||||||||||||||||||
| Entrez |
|
||||||||||||||||||||||||
| Ансамбль |
|
||||||||||||||||||||||||
| UniProt |
|
||||||||||||||||||||||||
| RefSeq (мРНК) |
|
|
|||||||||||||||||||||||
| RefSeq (белок) |
|
|
|||||||||||||||||||||||
| Расположение (UCSC) | Chr 14: 49.59 - 49.59 Мб | н / д | |||||||||||||||||||||||
| PubMed поиск | н / д | ||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||
| |||||||||||||||||||||||||



РНК частиц распознавания сигнала , (также известный как 7SL, 6S, эф или 4.5S РНК) является частью частицы распознавания сигнала (SRP) рибонуклеопротеинового комплекса. SRP распознает сигнальный пептид и связывается с рибосомой , останавливая синтез белка. SRP-рецептор - это белок, который встроен в мембрану и содержит трансмембранную пору . Когда комплекс SRP-рибосома связывается с рецептором SRP, SRP высвобождает рибосому и уносится прочь. Рибосома возобновляет синтез белка, но теперь белок движется через трансмембранную пору SRP-рецептора .
Таким образом, SRP направляет движение белков внутри клетки для связывания с трансмембранной порой, что позволяет белку пересекать мембрану туда, где это необходимо. РНК и белковые компоненты этого комплекса очень консервативны, но различаются в зависимости от царства жизни.
Общее семейство SINE Alu, вероятно, произошло из гена 7SL РНК после делеции центральной последовательности.
Эукариотической SRP состоит из 300 нуклеотидов 7S в РНК и шесть белков: SRPS 72, 68, 54, 19, 14 и 9. архейных SRP состоит из РНК и гомологов эукариотической SRP19 7S и SRP54 белков. 7S РНК эукариот и архей имеют очень похожие вторичные структуры.
У большинства бактерий SRP состоит из молекулы РНК (4.5S) и белка Ffh (гомолог эукариотического белка SRP54). Некоторые грамположительные бактерии (например, Bacillus subtilis ) имеют более длинную эукариотоподобную РНК SRP, которая включает домен Alu .
У эукариот и архей восемь спиральных элементов складываются в домены Alu и S, разделенные длинной линкерной областью. Полагают, что домен Alu опосредует функцию замедления удлинения пептидной цепи SRP. Универсально консервативная спираль, которая взаимодействует с M-доменом SRP54, опосредует распознавание сигнальной последовательности. Считается, что комплекс SRP19-спираль 6 участвует в сборке SRP и стабилизирует спираль 8 для SRP54. Связывание У людей есть три функциональных гена РНК SRP, которые удобно назвать RN7SL1, RN7SL2 и RN7SL3. В частности, известно, что геном человека содержит большое количество последовательности, связанной с РНК SRP, включая Alu-повторы .
Открытие
SRP РНК была впервые обнаружена в онкогенных РНК (окорна) вирусных частицах птиц и мышей . Впоследствии было обнаружено, что РНК SRP является стабильным компонентом неинфицированных клеток HeLa, где она связана с мембранными и полисомными фракциями. В 1980 году , клеточные биологи очищали из собачьей поджелудочной железы в 11S «белок распознавания сигнала» (также сокращенно единичными «SRP») , который способствовал транслокацию секреторных белков через мембрану из эндоплазматического ретикулума . Затем было обнаружено, что SRP содержит компонент РНК . Сравнение генов РНК SRP от разных видов показало, что спираль 8 РНК SRP является высококонсервативной во всех сферах жизни . Области около 5'- и 3'-концов SRP РНК млекопитающих подобны доминантному семейству Alu со средними повторяющимися последовательностями генома человека . Теперь понятно, что Alu ДНК произошла из РНК SRP путем вырезания центрального фрагмента РНК SRP (S) с последующей обратной транскрипцией и интеграцией во множество участков хромосом человека . РНК SRP были идентифицированы также в некоторых органеллах , например в пластидных SRP многих фотосинтезирующих организмов.
Транскрипция и обработка
РНК SRP эукариот транскрибируются из ДНК с помощью РНК-полимеразы III (Pol III). РНК-полимераза III также транскрибирует гены 5S рибосомальной РНК, тРНК , 7SK РНК и сплайсосомной РНК U6 . В промоторы генов SRP РНК человека включают в себя элементы , расположенные ниже по потоку от стартового сайта транскрипции. Промоторы РНК SRP растений содержат расположенный выше стимулирующий элемент (USE) и ТАТА-бокс . Гены дрожжевой РНК SRP имеют ТАТА-бокс и дополнительные внутригенные промоторные последовательности (называемые А- и В-блоками), которые играют роль в регуляции транскрипции гена SRP с помощью Pol III. В бактериях , гены организованы в опероне и транскрибируются РНК - полимераза . 5'-конец малого (4.5S) SRP РНК многих бактерий расщепляется с помощью РНКазы Р . Концы РНК SRP Bacillus subtilis процессируются РНКазой III . Пока интронов SRP РНК не обнаружено.
Функция

Ко-трансляционная транслокация
РНК SRP является неотъемлемой частью малого и большого домена SRP. Функция малого домена состоит в том, чтобы задерживать трансляцию белка до тех пор, пока связанный с рибосомами SRP не получит возможность связываться с мембранно-резидентным рецептором SRP (SR). Внутри большого домена РНК SRP заряженной сигнальным пептидом SRP способствует гидролизу двух молекул гуанозинтрифосфата (GTP). Эта реакция высвобождает SRP из рецептора SRP и рибосомы , позволяя трансляции продолжаться и белку поступать в транслокон . Белок пересекает мембрану совместно (во время трансляции) и входит в другой клеточный компартмент или внеклеточное пространство. В эукариот , мишень представляет собой мембрану из эндоплазматического ретикулума (ER). У архей SRP доставляет белки к плазматической мембране . У бактерий SRP в первую очередь включает белки во внутреннюю мембрану.
Посттрансляционный транспорт
SRP также участвует в сортировке белков после завершения их синтеза (посттрансляционная сортировка белков). У эукариот заякоренные в хвосте белки, обладающие гидрофобной последовательностью вставки на их С-конце, доставляются в эндоплазматический ретикулум (ER) с помощью SRP. Точно так же SRP помогает посттрансляционно импортировать ядерно-кодируемые белки на тилакоидную мембрану хлоропластов .
Состав

В 2005 г. номенклатура всех РНК SRP предлагала систему нумерации из 12 спиралей. Секции спирали обозначаются суффиксом строчной буквы (например, 5a). Вставки или «ответвления» спирали обозначены пунктирными номерами (например, 9.1 и 12.1).
РНК SRP охватывает широкий филогенетический спектр по размеру и количеству ее структурных особенностей (см. Примеры вторичной структуры SRP РНК ниже). Наименьшие функциональные РНК SRP были обнаружены у микоплазм и родственных видов. SRP-РНК Escherichia coli (также называемая 4.5S-РНК) состоит из 114 нуклеотидных остатков и образует стержневую петлю РНК . Грамположительные бактерии Bacillus зиЫШз кодирует больший 6S SRP РНК , которые напоминают архейные гомологи , но не хватает SRP РНК спирал 6. архейная SRP РНК обладает спиралями от 1 до 8, отсутствие спирали 7, и характеризуется третичной структурой , которая включает в себя апикальные петли спирали 3 и спирали 4. В SRP РНК эукариот отсутствует спираль 1 и они содержат спираль 7 переменного размера. Некоторые SRP-РНК простейших имеют уменьшенные спирали 3 и 4. SRP-РНК ascomycota имеют в целом уменьшенный малый домен и не имеют спиралей 3 и 4. Наибольшие известные на сегодняшний день SRP-РНК обнаружены у дрожжей ( Saccharomycetes ), которые приобрели спирали 9-12 как вставки в спираль 5, а также в удлиненную спираль 7. Семенные растения экспрессируют многочисленные сильно дивергентные РНК SRP.
Мотивы
Были идентифицированы четыре консервативных признака (мотива) (показаны на рисунке темно-серым цветом): (1) связывающий мотив SRP54, (2) мотив тетрапетли Helix 6 GNAR, (3) мотив 5e и (4) UGU (NR). мотив.
Привязка SRP54
Асимметричная петля между спиральными участками 8a и 8b и смежным участком 8b с парными основаниями является заметным свойством каждой SRP РНК. Спиральный участок 8b содержит пары оснований, отличных от Ватсона-Крика, которые способствуют образованию плоской малой бороздки в РНК, подходящей для связывания белка SRP54 (называемого у бактерий Ffh). Апикальная петля спирали 8 содержит четыре, пять, шесть или остатков, в зависимости от вида . Он имеет высококонсервативный гуанозин в качестве первого и аденозин в качестве последнего остатка петли. Эта особенность требуется для взаимодействия с третьим аденозиновым остатком тетрапетлевой спирали 6 GNAR.
Тетралуп Helix 6 GNAR
SRP РНК эукариот и архей имеют тетрапетлю GNAR (N - для любого нуклеотида , R - для пурина ) в спирали 6. Его консервативный аденозиновый остаток важен для связывания белка SRP19. Этот аденозин вступает в третичное взаимодействие с другим остатком аденозина, расположенным в апикальной петле спирали 8.
5e
11 нуклеотидов мотива 5e образуют четыре пары оснований, которые прерываются петлей из трех нуклеотидов . У эукариот первым нуклеотидом петли является аденозин, который необходим для связывания белка SRP72.
UGU (NR)
Мотив UGU (NR) соединяет спирали 3 и 4 в малом (Alu) домене SRP. Грибковые РНК SRP, лишенные спиралей 3 и 4, содержат мотив внутри петли спирали 2. Это важно для связывания гетеродимера белка SRP9 / 14 как части разворота РНК .
Вторичный
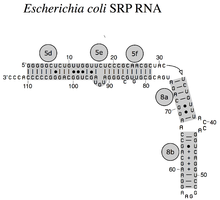
Бактериальная РНК SRP (4.5S РНК) из E. coli

Бактериальная РНК SRP (6S РНК) из Bacillus subtilis
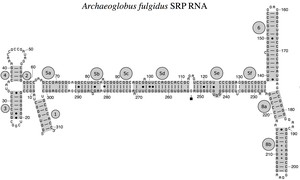
Архейная SRP РНК Archaeoglobus fulgidus

SRP РНК эукариотических протистов из Trypanosoma brucei

SRP РНК эукариотических дрожжей из Saccharomyces cerevisiae





Третичный
| SRP РНК | |
|---|---|
| Идентификаторы | |
| Рфам | CL00003 |
| Прочие данные | |
| Структуры PDB | PDBe 2IY3 , 1Z43 , 1RY1 , 1QZW , 1MFQ , 1L9A , 1LNG , 1JID , 1E8S , 1E8O , 1DUL , 1DUH , 1D4R , 28SR , 28SP |
Рентгеновская кристаллография , ядерный магнитный резонанс (ЯМР) и криоэлектронная микроскопия (крио-ЭМ) использовались для определения молекулярной структуры частей РНК SRP различных видов . Доступные структуры PDB показывают, что молекула РНК либо свободна. или когда он связан с одним или несколькими белками SRP .

SRP19-7S.S SRP РНК-комплекс из M. jannaschii

S-домен человеческого SRP


Связывающие белки
Один или несколько белков SRP связываются с РНК SRP для сборки функционального SRP. Белки SRP названы в соответствии с их приблизительной молекулярной массой, измеренной в килодальтонах . Большинство бактерий Политики SRP состоят из SRP РНК и SRP54 (также названный FFH для « F ifty- ф наш час УДОСТОВЕРЕ»). Архейных SRP содержит белки SRP54 и SRP19. У эукариот РНК SRP комбинируется с импортированными белками SRP SRP9 / 14, SRP19 и SRP68 / 72 в области ядрышка . Этот пре-SRP транспортируется в цитозоль, где связывается с белком SRP54. В молекулярные структуры свободной или SRP РНК-связанных белков SRP9 / 14, SRP19 или SRP54 известны с высоким разрешением.
SRP9 и SRP14
SRP9 и SRP14 структурно родственны и образуют гетеродимер SRP9 / 14, который связывается с SRP РНК малого (Alu) домена. SRP дрожжей лишен SRP9 и содержит структурно связанный связывающий белок SRP21. SRP14 дрожжей образует гомодимеры в кристаллах и не связывает Alu. SRP9 / 14 отсутствует в SRP трипаносомы, которая вместо этого имеет молекулу, подобную тРНК .
SRP19
SRP19 обнаружен в SRP эукариот и архей . Его основная роль заключается в подготовке РНК SRP для связывания SRP54, SRP68 и SRP72 путем правильной организации спиралей 6 и 8 РНК SRP. SRP дрожжей содержит Sec65p, более крупный гомолог SRP19.
SRP54
Белок SRP54 (называемый у бактерий Ffh ) является важным компонентом каждого SRP. Он состоит из трех функциональных доменов : N-концевого (N) домена, домена GTPase (G) и домена, богатого метионином (M).
SRP68 и SRP72
Белки SRP68 и SRP72 являются структурно неродственными составляющими большого домена эукариотического SRP. Они образуют стабильный гетеродимер SRP68 / 72. Было показано, что около одной трети человеческого белка SRP68 связывается с РНК SRP. Относительно небольшая область, расположенная рядом с С-концом SRP72, связывается с мотивом РНК 5e SRP.
использованная литература
дальнейшее чтение
- Kuglstatter A, Oubridge C, Nagai K (октябрь 2002 г.). «Вызванные структурные изменения 7SL РНК во время сборки человеческих сигнальных частиц». Структурная биология природы . 9 (10): 740–744. DOI : 10.1038 / nsb843 . PMID 12244299 . S2CID 9543041 .
- Ван Т., Тиан Ч., Чжан В., Ло К., Саркис П. Т., Ю Л, Лю Б., Ю Й, Ю. XF (декабрь 2007 г.). «7SL РНК опосредует упаковку вириона противовирусной цитидиндезаминазы APOBEC3G» . Журнал вирусологии . 81 (23): 13112–13124. DOI : 10,1128 / JVI.00892-07 . PMC 2169093 . PMID 17881443 .
- Уллу Э., Вайнер А.М. (декабрь 1984 г.). «Человеческие гены и псевдогены для компонента 7SL РНК частицы распознавания сигнала» . Журнал EMBO . 3 (13): 3303–3310. DOI : 10.1002 / j.1460-2075.1984.tb02294.x . PMC 557853 . PMID 6084597 .
- Энглерт М., Фелис М., Юнкер В., Байер Х (декабрь 2004 г.). «Новые вышестоящие и внутригенные элементы контроля для зависимой от РНК-полимеразы III транскрипции генов 7SL РНК человека». Биохимия . 86 (12): 867–874. DOI : 10.1016 / j.biochi.2004.10.012 . PMID 15667936 .
- Обридж К., Куглштаттер А., Джовин Л., Нагаи К. (июнь 2002 г.). «Кристаллическая структура SRP19 в комплексе с S доменом SRP РНК и ее значение для сборки сигнальной частицы распознавания» . Молекулярная клетка . 9 (6): 1251–1261. DOI : 10.1016 / S1097-2765 (02) 00530-0 . PMID 12086622 .
- Ван Т., Тиан Ч., Чжан В., Саркис П. Т., Ю XF (январь 2008 г.). «Для упаковки вириона APOBEC3F требуется взаимодействие с 7SL РНК, но не с геномной РНК ВИЧ-1 или Р-тельцами». Журнал молекулярной биологии . 375 (4): 1098–1112. DOI : 10.1016 / j.jmb.2007.11.017 . PMID 18067920 .
внешние ссылки
- База данных SRP (SRPDB): выравнивание РНК SRP и связанных белков, вторичных структур SRP РНК и трехмерных моделей.
- Запись Rfam для РНК частицы распознавания сигнала типа Metazoan
- Запись Rfam для РНК бактериальной частицы, распознающей малый сигнал
- Запись Rfam для РНК бактериальной частицы с большим сигналом распознавания
- Запись Rfam для РНК частицы, распознающей сигнал грибка
- Запись Rfam для РНК частицы распознавания сигнала растений
- Запись Rfam для РНК частицы распознавания сигнала простейшими
- Запись Rfam для РНК частицы распознавания сигнала архей
- Фильм о распознавании сигналов Dnatube Particle