
Список биофизически важных структур макромолекулярных кристаллов - List of biophysically important macromolecular crystal structures

Кристаллические структуры молекул белков и нуклеиновых кислот и их комплексов занимают центральное место в практике большинства областей биофизики и сформировали большую часть того, что мы понимаем с научной точки зрения на уровне детализации атома биологии. Их важность подчеркивается Организацией Объединенных Наций, объявляющей 2014 год Международным годом кристаллографии , как 100-летие Нобелевской премии Макса фон Лауэ 1914 года за открытие дифракции рентгеновских лучей на кристаллах. Этот хронологический список биофизически значимых структур белков и нуклеиновых кислот частично основан на обзоре, опубликованном в Biophysical Journal . Список включает в себя все первые дюжины отдельных структур, тех, которые открыли новые горизонты в предмете или методе, и те, которые стали модельными системами для работы в будущих биофизических областях исследований.
Миоглобин


1960 - Миоглобин был самой первой кристаллической структурой белковой молекулы с высоким разрешением. Миоглобин удерживает железосодержащую гемовую группу, которая обратимо связывает кислород для использования в мышечных волокнах, и эти первые кристаллы были миоглобином кашалота , мышцы которого нуждаются в обильном хранении кислорода для глубоких погружений. Трехмерная структура миоглобина состоит из 8 альфа-спиралей , и кристаллическая структура показала, что их конформация была правосторонней и очень близко соответствовала геометрии, предложенной Линусом Полингом , с 3,6 остатками на оборот и водородными связями основной цепи из пептида. NH одного остатка к пептиду CO остатка i + 4. Миоглобин является модельной системой для многих типов биофизических исследований, особенно связанных с процессом связывания небольших лигандов, таких как кислород и окись углерода .
Гемоглобин
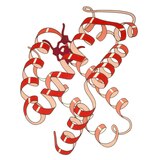

1960 - Кристаллическая структура гемоглобина показала тетрамер из двух связанных типов цепей и была решена с гораздо более низким разрешением, чем мономерный миоглобин, но он явно имел ту же базовую 8-спиральную архитектуру (теперь называемую «глобиновой складкой»). Дальнейшие кристаллические структуры гемоглобина с более высоким разрешением [PDB 1MHB, 1DHB) вскоре показали сопряженное изменение как локальной, так и четвертичной конформации между кислородным и дезокси-состояниями гемоглобина, что объясняет кооперативность связывания кислорода в крови и аллостерический эффект таких факторов, как как pH и DPG . На протяжении десятилетий гемоглобин был основным примером концепции аллостерии, а также предметом интенсивных исследований и дискуссий по аллостерии. В 1909 г. кристаллы гемоглобина более 100 видов использовались для соотнесения таксономии с молекулярными свойствами. Эту книгу процитировал Перуц в отчете о кристаллах конского гемоглобина за 1938 год, с которого началась его длинная сага о кристаллической структуре. Кристаллы гемоглобина плеохроичны - темно-красные в двух направлениях и бледно-красные в третьем - из-за ориентации гемов, а яркая полоса Соре гем- порфириновых групп используется в спектроскопическом анализе связывания гемоглобинового лиганда.

Лизоцим курино-яичного белка
1965 - Курочка-яичный белок лизоцим (PDB файл 1lyz). была первой кристаллической структурой фермента (он расщепляет небольшие углеводы на простые сахара), использованной для ранних исследований механизма фермента. Он содержал бета-лист (антипараллельный), а также спирали, а также был первой макромолекулярной структурой, у которой были уточнены координаты атомов (в реальном пространстве). Исходный материал для приготовления можно купить в продуктовом магазине, а лизоцим куриных яиц очень легко кристаллизуется во многих различных пространственных группах ; это любимый тестовый образец для новых кристаллографических экспериментов и инструментов. Недавними примерами являются нанокристаллы лизоцима для сбора данных лазера на свободных электронах и микрокристаллы для дифракции микроэлектронов.

Рибонуклеаза
1967 - Рибонуклеаза A (файл PDB 2RSA) представляет собой фермент, расщепляющий РНК, стабилизированный 4 дисульфидными связями. Он был использован в основополагающих исследованиях Анфинсена по сворачиванию белка, которые привели к концепции, что трехмерная структура белка определяется его аминокислотной последовательностью. Рибонуклеаза S , расщепленная двухкомпонентная форма, изученная Фредом Ричардсом , также была ферментативно активной, имела почти идентичную кристаллическую структуру (файл PDB 1RNS) и, как было показано, была каталитически активна даже в кристалле, что помогло развеять сомнения относительно актуальности кристаллических структур белка к биологической функции.

Сериновые протеазы
1967 - Сериновые протеазы представляют собой исторически очень важную группу структур ферментов, поскольку в совокупности они освещают каталитический механизм (в их случае «каталитической триадой» Ser-His-Asp), основу различных субстратных специфичностей и механизм активации. с помощью которого контролируемое ферментативное расщепление закапывает новый конец цепи, чтобы правильно перестроить активный сайт. Ранние кристаллические структуры включали химотрипсин (файл PDB 2CHA), химотрипсиноген (файл PDB 1CHG), трипсин (файл PDB 1PTN) и эластазу (файл PDB 1EST). Они также были первыми белковыми структурами, в которых обнаружены два почти идентичных домена, предположительно связанных дупликацией генов . Одной из причин их широкого использования в качестве примеров в учебниках и аудиториях была система нумерации вставных кодов (которую ненавидят все компьютерные программисты), которая сделала Ser195 и His57 последовательными и запоминающимися, несмотря на различия в последовательностях, специфичных для белков.
Папаин
1968 - Папаин


Карбоксипептидаза
1969 - Карбоксипептидаза А - металлопротеиназа цинка . Его кристаллическая структура (файл PDB 1CPA) показала первую параллельную бета-структуру: большой скрученный центральный лист из 8 нитей с Zn в активном центре, расположенным на С-конце средних цепей, и лист, фланкированный с обеих сторон альфа-спирали. Это экзопептидаза, которая отщепляет пептиды или белки от карбокси-конца, а не внутри последовательности. Позже был обнаружен небольшой белковый ингибитор карбоксипептидазы (файл PDB 4CPA), который механически останавливает катализ, представляя его С-концевой конец просто торчащим из между кольцом дисульфидных связей с плотной структурой позади него, предотвращая всасывание фермента в цепи. мимо первого остатка.

Субтилизин
1969 - Субтилизин (файл PDB 1sbt) был вторым типом сериновой протеазы с почти идентичным активным сайтом ферментам семейства трипсина, но с совершенно другой общей структурой. Это дало первое представление о конвергентной эволюции на атомном уровне. Позже интенсивное мутационное исследование субтилизина задокументировало эффекты всех 19 других аминокислот в каждом отдельном положении.
Лактатдегидрогеназа
1970 - Лактатдегидрогеназа

Ингибитор трипсина
1970 - Базовый ингибитор трипсина поджелудочной железы , или BPTI (файл PDB 2pti), представляет собой небольшой, очень стабильный белок, который представляет собой высокопродуктивную модельную систему для изучения сверхпрочного связывания, образования дисульфидной связи (SS), сворачивания белка , молекулярной стабильности. за счет аминокислотных мутаций или обмена водород-дейтерий и быстрой локальной динамики с помощью ЯМР . Биологически BPTI связывает и ингибирует трипсин, пока он хранится в поджелудочной железе , позволяя активировать переваривание белка только после того, как трипсин попадает в желудок.
Рубредоксин
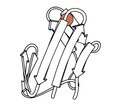
1970 - Рубредоксин (файл PDB 2rxn) был первой решенной редокс-структурой, минималистским белком с железом, связанным с 4 боковыми цепями цис из 2 петель на вершине β-шпилек. Он дифрагировал до 1,2 Å, что сделало возможным первое уточнение белка в обратном пространстве (4,5rxn). [NB: остерегайтесь 4rxn, сделано без геометрических ограничений!] Архейские рубредоксины составляют многие из небольших структур с самым высоким разрешением в PDB.


Инсулин
1971 - Инсулин (файл PDB 1INS) - это гормон, имеющий центральное значение для метаболизма сахара и накопления жира и важный при таких заболеваниях человека, как ожирение и диабет . Он биофизически примечателен своим связыванием Zn, равновесием между состояниями мономера, димера и гексамера, его способностью образовывать кристаллы in vivo и его синтезом в виде более длинной «про» формы, которая затем расщепляется и складывается в виде активного 2- цепь, SS-связанный мономер. Инсулин оказался успешным в рамках программы НАСА по выращиванию кристаллов на космическом шаттле , в результате чего были получены препараты очень однородных крошечных кристаллов для контролируемой дозировки.
Стафилококковая нуклеаза
1971 - стафилококковая нуклеаза
Цитохром с
1971 - Цитохром С
Лизоцим фага Т4
1974 - лизоцим фага Т4
Иммуноглобулины
1974 - Иммуноглобулины
Супероксиддисмутаза
1975 - Cu, Zn Супероксиддисмутаза
Передача РНК
1976 - Трансфер РНК

Триозофосфат изомераза
1976 - Триозофосфат изомераза
Пепсиноподобные аспарагиновые протеазы
1976 - Ризопуспепсин
1976 - Эндотиапепсин
1976 - Пенициллопепсин
Более поздние постройки (с 1978 г.)
1978 - Икосаэдрический вирус
1981 - Додекамер ДНК B-формы Дикерсона
1981 - Крамбин
1985 - Кальмодулин
1985 - ДНК-полимераза

1985 - Центр фотосинтетических реакций Пары бактериохлорофиллов (зеленые) внутри мембраны улавливают энергию солнечного света, затем перемещаются по многим ступеням, чтобы стать доступными в гемовых группах (красный) в модуле цитохрома-С вверху. Это была первая кристаллическая структура, решенная для мембранного белка, веха, отмеченная Нобелевской премией Хартмуту Мишелю, Гансу Дайзенхоферу и Роберту Хуберу.
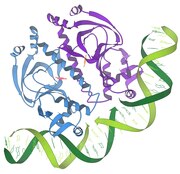
1986 - Взаимодействие репрессора / ДНК
1987 - Большой комплекс гистосовместимости '
1987 - Убиквитин
1987 - белок ROP
1989 - протеаза ВИЧ-1
1990 - Бактериородопсин
1991 - спиральная катушка GCN4
1991 - обратная транскриптаза ВИЧ-1
1993 - Бета спираль из пектата лиазы
1994 - Коллаген
1994 - Комплекс Barnase / barstar
1994 - АТФаза F1
1995 - Гетеротримерные G-белки
1996 - Зеленый флуоресцентный белок

1996 - Моторный белок кинезин
1997 - Шаперон GroEL / ES
1997 - Нуклеосома
1998 - Самосплайсинговый интрон I группы

1998 - ДНК-топоизомеразы выполняют биологически важную и необходимую работу по распутыванию цепей ДНК или спиралей, которые переплетаются друг с другом или слишком сильно перекручиваются во время нормальных клеточных процессов, таких как транскрипция генетической информации.
1998 - димер тубулина альфа / бета
1998 - Калиевый канал
1998 - перекресток Холлидей
2000 - Рибосомы являются центральной частью биологии и биофизики, которая впервые стала структурно доступной в 2000 году.

2000 - ААА + АТФаза
2002 - Анкирин повторяет
2003 - ТОП7 белкового дизайна
2004 - Цианобактериальные белки циркадных часов
2004 - Рибосвич
2006 - Экзосома человека

2007 - рецептор, связанный с G-белком

2009 - Частица Убежища - это новое интригующее открытие большой полой частицы, часто встречающейся в клетках, с несколькими различными предположениями о ее возможной биологической функции. Кристаллические структуры (файлы PDB 2zuo, 2zv4, 2zv5 и 4hl8) показывают, что каждая половина хранилища состоит из 39 копий длинного 12-доменного белка, которые вращаются вместе, образуя оболочку. Беспорядок на самом верхнем и нижнем концах предполагает отверстия для возможного доступа внутрь хранилища.
