
Формирование парка динозавров - Dinosaur Park Formation
| Формирование парка динозавров Стратиграфический диапазон : поздний мел , кампан ,76,9–75,8 млн лет | |
|---|---|
 Формирование парка динозавров, обнаженное вдоль реки Ред-Дир в провинциальном парке динозавров , на юго-востоке Альберты, Канада.
| |
| Тип | Геологическое образование |
| Единица | Belly River Group |
| Лежит в основе | Формация Медвежьей Лапы |
| Перекрывает | Формация Олдмана |
| Литология | |
| Начальный |
Песчаник (нижний), аргиллиты и алевролиты (верхний) |
| Другой | Бентонит и уголь |
| Место нахождения | |
| Координаты | 49 ° 12'N 110 ° 24'W / 49.2 ° N 110.4 ° W Координаты : 49.2 ° N 110.4 ° W49 ° 12'N 110 ° 24'W / |
| Примерные палеокоординаты | 56 ° 24'N 75 ° 48'W / 56,4 ° с. Ш. 75,8 ° з. |
| Область |
|
| Страна |
|
| Степень | Западноканадский осадочный бассейн |
| Тип раздела | |
| Названный для | Провинциальный парк динозавров |
| Названный | Эберт Д.А. и Хамблин А.П. |
| Год определен | 1993 г. |
 Формирование парка динозавров (Канада)
 Формация парка динозавров (Альберта)
| |
Парк динозавров Формирование является самым верхним членом Belly River Group (также известный как River Group Джудит ), крупный геологический блок в южной части провинции Альберта . Он был отложен на кампанском этапе позднего мела , примерно между 76,9 и 75,8 миллионами лет назад. Он был отложен на аллювиальных и прибрежных равнинах и ограничен неморской формацией Олдман под ней и морской формацией Медвежья Лапа над ней.
Формация парка динозавров содержит плотные скопления скелетов динозавров , как сочлененных, так и разрозненных, которые часто встречаются вместе с сохранившимися остатками мягких тканей. Остатки других животных, таких как рыбы , черепахи и крокодилы , а также остатки растений также в изобилии. Формация была названа в честь Провинциального парка динозавров , объекта Всемирного наследия ЮНЕСКО, где формация хорошо видна в бесплодных землях , примыкающих к реке Ред-Дир .
Геологическая обстановка

Формация «Парк динозавров» состоит из отложений, образовавшихся в результате эрозии гор на западе. Он был отложен на аллювиальной прибрежной равнине речными системами, которые текли на восток и юго-восток в море Медвежьей Лапы, большое внутреннее море, которое было частью Западного внутреннего морского пути . Это море постепенно затопило прилегающую прибрежную равнину, осаждение морских сланцев на формировании BEARPAW на вершине Dinosaur Park свиты.
Формация парка динозавров имеет толщину около 70 метров (230 футов) в парке динозавров. Нижняя часть формации залегает в средах русла рек и состоит в основном из мелкозернистых и среднезернистых песчаников с пересеченными пластами . Верхняя часть, которая была отложена в условиях надбровья и поймы , состоит в основном из массивных или слоистых, богатых органикой аргиллитов с обильными следами корней и тонких пластов бентонита . Угольная зона Летбридж, состоящая из нескольких пластов низкосортного угля с прослоями аргиллитов и алевролитов , отмечает кровлю формации.
Отложения формации «Парк динозавров» аналогичны отложениям нижележащей формации Олдман, и изначально они были включены в эту формацию. Однако эти две формации разделены региональным несоответствием и отличаются петрографическими и седиментологическими различиями. Кроме того, сочлененные останки скелета и костяные пласты редки в формации Олдман, но многочисленны в формации парка динозавров.
Биостратиграфия
Формация Парка динозавров может быть разделена как минимум на две отдельные фауны. Нижняя часть свиты характеризуется обилием коритозавров и центрозавров . Эта группа видов заменяется на более высоких ступенях формацией другой орнитисхийской фауной, для которой характерно присутствие ламбеозавров и стиракозавров . Появление нескольких новых, редких видов орнитисхиев на самой вершине формации может указывать на то, что третья особая фауна заменила вторую во время перехода к более молодым, не относящимся к Парку динозавров отложениям, в то время как внутреннее море выходит на сушу. , но здесь осталось меньше. Безымянный пахиринозавр , Vagaceratops irvinensis и Lambeosaurus magnicristatus могут быть более распространены в этой третьей фауне.
Приведенная ниже временная шкала следует за синтезом, представленным Фаулером (2016) с дополнительной информацией от Arbor et al. 2009 г., Evans et al. 2009, и Penkalski, 2013. Зоны скопления крупных травоядных животных (MAZ) соответствуют данным, представленным Mallon et al. , 2012.

Амфибии
В свите обнаружены останки следующих земноводных:
Albanerpetontidae (вымершие амфибии, похожие на саламандр )
Хвостатые ( саламандры )
- Habrosaurus prodilatus
- Лиссерпетон
- Опистотритон кайи
- Скаферпетон тектум
- безымянный хвостатан
- 2 неопределенных хвостатых
- 2 безымянных салиентана
- Tyrrellbatrachus brinkmani
- Hensonbatrachus kermiti
Динозавры
В свите обнаружены останки следующих динозавров:
Орнитисхианы
В свите обнаружены остатки следующих орнитисхий:
Анкилозавры
| Анкилозавры из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Анодонтозавр | А. inceptus | Средний, 75.6 млн. Назад | [Два] черепа с зубами, нижней челюстью, частично подготовленным скелетом, шейными полукольцами и остеодермами. | Ankylosaurine ankylosaurid |
 |
|
| Диоплозавр | D. acutosquameus | Нижний, 76,5 млн. Назад | Частичный череп и скелет, включая таз, хвост и заднюю конечность с стопой и остеодермами. | Анкилозаврин анкилозаврид |
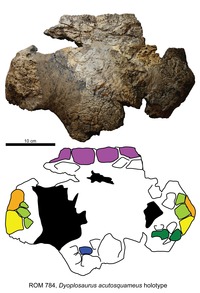 |
|
| Эдмонтония | E. rugosidens | Нижний, 76,5-75,9 млн лет назад | Частичный скелет, включающий череп, спинные позвонки, проксимальный, дистальный каудальный отдел, ребра, плечевую кость, локтевую кость, лучевую кость, кисть, фрагменты таза, большеберцовой кости, малоберцовой кости?, Остеодермы; передняя половина сочлененного скелета с остеодермами in situ и парные первые медиальные щитки. | Nodosaurine nodosaurid также известен из свиты подковы Каньона и две медицины формирования |
 |
|
| Евоплоцефал | E. пачки | От нижнего к среднему, ~ 76,4-75,6 млн лет назад | [Четыре] черепа, нижней челюсти, шейных позвонков, спинных позвонков, ребер, лопаток, плечевой кости, лучевой кости, локтевой кости, пястных костей, фаланги, подвздошной кости, седалищной кости, бедренной кости, голени, частичной стопы, крестца, шейных полуколец и остеодермы. | Анкилозаврин анкилозаврид |
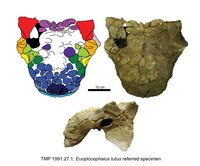 |
|
| Паноплозавр | P. mirus | Средний, 75.6 млн. Назад | Череп с нижней челюстью, изолированные зубы, шейные позвонки, спинные позвонки, крестцовые позвонки, шейные ребра, задние ребра, скапулокоракоид, плечевая кость, кисть, большеберцовая кость, малоберцовая кость, окостеневшая внутренняя пластинка, пара окостеневших мечевидных отростков, язвы и кости in situ . | Нодозаврид нодозавринов |
 |
|
| Платипельта | P. coombsi | Нижний, 77,5-76,5 млн лет назад | Хорошо сохранившийся череп, нижние челюсти, зубы, шейные и спинные позвонки, ребра, таз в целом, оба скапулокоракоида, плечевые и лучевые кости, оба шейных полукольца и остеодермы. | Анкилозаврин анкилозаврид |
 |
|
| Сколозавр | С. катлери | Нижний, 76,5 млн лет назад или более | Почти полный скелет, череп, шейный, спинной и хвостовой позвонки, ребра, лопатка, коракоид, плечевые кости, радиусы, подвздошная кость, седалищная кость, бедренная кость, большеберцовая кость, малоберцовая кость, [одно] шейное полукольцо и остеодермы. | Анкилозаврин анкилозаврид недолго считался синонимом Euoplocephalus . Возможно, он пришел из верхних слоев нижележащей формации Олдмана . |
 |
|
| S. thronus | Верхний, 75 млн назад | Частичный скелет, включающий череп, спинные позвонки, полный синсакрум, крестцовые ребра, хвостовые позвонки, лопатку, частичные подвздошные кости, плечевую кость, шейные полукольца, остеодермы и отпечатки кожи. | Анкилозаврин анкилозаврид | |||
Цератопсы
Из формации был извлечен безымянный таксон, похожий на пахиринозавра.
| Цератопсы из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Центрозавр | C. apertus | Middle, 76.2-75.5Ma назад | «[Пятнадцать] черепов, несколько скелетов, все взрослые; обильный материал костного ложа с редкими молодыми и половозрелыми особями». C. nasicornis может быть синонимом. | Центрозаврины ceratopsid |
 |
|
| Хасмозавр | C. belli | Средний, 76-75,5 млн. Назад | «[Двенадцать] черепов, несколько скелетов». | Хасмозаврины ceratopsid |
 |
|
| К. Расселли | Нижний, 76.5-76Ма назад | «[Шесть] полных или частичных черепов». | ||||
| Меркурицератопс | М. близнецы | Нижний, ~ 77 млн. Назад | «одна апоморфная чешуя» | Хасмозаврин цератопсид |
 |
|
| Моноклониус | М. lowei | Сомнительные центрозаврины ceratopsid. Возможно, это синоним Центрозавра . | ||||
| Пентацератопс | P. aquilonius | Самый верхний, 74,8 МА | два фрагмента оборки | Сомнительный хазмозаврин цератопсид, который может принадлежать к тому же виду, что и Spiclypeus shipporum . |
 |
|
| Спинопс | С. штернбергорум | Нижний, 76,5 мА | «частичная теменная кость, частичная зубная кость, неидентифицируемые фрагменты конечностей, частичный череп и частичный правый чешуйчатый». | Центрозаврин цератопсид. На самом деле он может быть из верхней формации Олдмана. |
 |
|
| Стиракозавр | S. albertensis | Upper, 75.5-75.2Ma назад | «[Два] черепа, [три] скелета, дополнительный материал в костных ложах». | Центрозаврин цератопсид |
 |
|
| ЮНЕСКОцератопс | U. koppelhusae | Неполная нижняя челюсть | Leptoceratopsid полагают, были от одного до двух метров в длину и не менее 91 кг. Его зубы были самыми круглыми из всех лептоцератопсид. | |||
| Вагацератопс | V. irvinensis | Upper, 75Мнн назад | «[Три] черепа, скелет без хвоста». | Вид хасмозавров цератопсид, ранее классифицированный как вид хасмозавров . |
 |
|
Орнитоподы
По крайней мере, один экземпляр неопределенного гипсилофодонта был извлечен из формации.
В обзоре яичной скорлупы и материала вылупившихся гадрозавров из формации «Парк динозавров» в 2001 году Даррен Х. Танке и М.К. Бретт-Сурман пришли к выводу, что гадрозавры гнездились как на древних возвышенностях, так и на низинах в среде осадконакопления формации. Горные места гнездования могли быть предпочтительны для менее распространенных гадрозавров, таких как брахилофозавр или паразауролоф . Однако авторам не удалось определить, какие именно факторы повлияли на выбор места гнездования у гадрозавров формации. Они предположили, что поведение, диета, состояние почвы и конкуренция между видами динозавров потенциально влияют на то, где гнездятся гадрозавры.
Сообщалось о субсантиметровых фрагментах яичной скорлупы гадрозавров с текстурой гальки из формации «Парк динозавров». Эта яичная скорлупа похожа на яичную скорлупу гадрозавра в Devil's Coulee в южной Альберте, а также на яичные формации Two Medicine и Judith River в Монтане, США. В настоящее время яичная скорлупа динозавров очень редко встречается в формации Парка динозавров и встречается только в двух разных местах обнаружения ископаемых. Эти участки отличаются большим количеством моллюсков- писидидов и других менее распространенных беспозвоночных с панцирем, таких как моллюски- юниониды и улитки. Эта ассоциация не случайна, поскольку панцири беспозвоночных медленно растворяются и выделяют достаточно основного карбоната кальция для защиты яичной скорлупы от встречающихся в природе кислот, которые в противном случае растворили бы их и предотвратили бы окаменелость.
В отличие от окаменелостей яичной скорлупы, останки очень молодых гадрозавров встречаются довольно часто. Даррен Танке заметил, что опытный коллекционер может обнаружить несколько особей молодых гадрозавров за один день. Наиболее часто встречающиеся остатки молодых гадрозавры в Dinosaur Park свиты являются dentaries , кости из конечностей и ног, а также позвоночный Centra . На материале не было практически никакого истирания, которое могло бы возникнуть в результате транспортировки, а это означает, что окаменелости были захоронены недалеко от места их происхождения. Костные слои 23, 28, 47 и 50 являются продуктивными источниками остатков молодых гадрозавров в формации, особенно костного ложа 50. Кости молодых гадрозавров и ископаемые фрагменты яичной скорлупы не сохранились вместе друг с другом, несмотря на то, что оба они присутствуют в формирование.
| Орнитоподы из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Коритозавр | C. casuarius | Нижнее-Среднее, 76,5–75,5 млн лет назад | «Приблизительно [десять] сочлененных черепов и связанных с ними посткраний, [от десяти до пятнадцати] сочлененных черепов, отдельные элементы черепа, от юношеского до взрослого». | Lambeosaurin lambeosaurine гадрозавр |
 |
|
| Грипозавр | G. notabilis | Нижний, 76.2-76Ма назад | «Примерно [десять] полных черепов, [двенадцать] фрагментарных черепов, связанных посткранией». | Kritosaurin saurolophine гадрозавр |
 |
|
| Ламбеозавр | L. lambei | Upper, 75.5-75Ма назад | «Приблизительно [семь] сочлененных черепов с соответствующими посткраниями, [возможно, десять] сочлененных черепов, отдельные элементы черепа, от юношеского до взрослого». | |||
| L. magnicristatus | Формация Upper / Bearpaw , 74,8 млн назад | «[Два] полных черепа, один с сочлененной посткранией». |
 |
|||
| Паразауролоф | П. уокери | Нижний, 76,5-75,3 млн. Лет назад | «Полный череп и посткраниальный скелет». | Parasaurolophin lambeosaurine гадрозавр. |
 |
|
| Прозавролоф | P. maximus | Верхний, 75,5 - 74,8 млн лет | «[От двадцати до двадцати пяти] человек, включая не менее [семи] сочлененных черепов и связанных с ними посткраний». | Saurolophin saurolophine гадрозавр |
 |
|
Пахицефалозавры
| Пахицефалозавры из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Foraminacephale | F. brevis | Также присутствует в формации Олдмана. | Фронтопаретальный купол, различные другие фрагменты черепа, включая подростковый и подростковый материал | Когда-то считалось разновидностью стегоцерас |

|
|
| Гравитолус | G. albertae | «Лобно-теменный купол». |
 |
|||
| Hanssuesia | Х. штернберги | Нижний, также присутствует в формациях Олдман и Джудит Ривер. |
 |
|||
| Стегоцерасы | С. валидум | Образцы, включая лобно-теменный купол. |
 |
|||
Теропод
В формации «Парк динозавров» маленькие тероподы встречаются редко из-за того, что их тонкостенные кости часто ломаются или плохо сохраняются. Маленькие кости маленьких теропод, на которые охотились более крупные, могли быть проглочены целиком и переварены. В этом контексте особенно ценным было открытие небольшого динозавра-теропод с сохранившимися следами зубов. Возможные неопределенные останки авимимидов известны из формации.
Орнитомимиды
| Орнитомимиды из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Орнитомим | O. sp. | Тип образца | Ornithomimid , возможно , один из видов Struthiomimus . |
 |
||
| Qiupalong | Q. sp. | Несколько экземпляров | Ornithomimid , возможно , излучение этого рода в Азии. |
 |
||
| Ратифицирует | Р. уклоняется | Тип образца | Орнитомимид, ранее представлявший собой Struthiomimus . |
 |
||
Овирапторозавры
Цветовой ключ
|
Примечания Неопределенные или предварительные таксоны выделены мелким шрифтом ; |
| Овирапторозавры из формации парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Caenagnathus | C. collinsi | Нижняя челюсть, типовой образец | Ценагнатид, который по размеру соперничал с Анзу . |
 |
||
| Хиростеноты | C. pergracilis | Несколько фрагментарных экземпляров, типовой образец | Ценагнатид среднего размера. |
 |
||
| Citipes | C. elegans | Несколько фрагментарных экземпляров, типовой образец | Самый мелкий ценагнатид из свиты. |
 |
||
| Макрофалангией | M. canadensis | Младший синоним Chirostenotes pergracilis | ||||
Паравианцы
Новый таксон троодонтид, основанный исключительно на зубах, известен из верхней части формации.
| Паравианцы из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| ср. Бапторнис | Неопределенный | Hesperornithine птица | ||||
| ср. Цимолоптерикс | Неопределенный | Частичный коракоид | Возможная рогатая птица | |||
| Дромеозавр | D. albertensis | Несколько экземпляров и зубов, типовой образец | Дромеозаврид |
 |
||
| Геспероних | Х. elizabethae | Тазобедренные кости и частичные пальцы и когти, типовой образец | Microraptorine dromaeosaur, также находится в Oldman формирования |
 |
||
| Латенивенатрикс | L. mcmasterae | Тазобедренные кости, таз, фрагменты черепа, типовой препарат | Большой троодонтид размером 3–3,5 м (9,8–11,5 футов). |
.png) |
||
| ср. Палинтроп | Безымянный | Неполные плечевые пояса | Ambiortiform птица | |||
| ср. Парониходон | ср. P. lacustris | Зубы | Неопределенный манирапторан, также найденный в реке Джудит. | |||
| ср. Пектинодон | Неопределенный | Зубы | Троодонт | |||
| Полиодонтозавр | P. grandis | Стоматологический образец | Nomen dubium. Возможно синоним латенивенатрикс. | |||
| Ричардэстезия | Р. гилмори | Нижняя челюсть , типовой образец | Дромеозаврид | |||
| R. равнобедренный | Зубы | |||||
| Saurornitholestes | S. langstoni | Неполный скелет и зубы, типовой экземпляр . Нижнечелюстной называют Saurornitholestes было обнаружено , что сохранились следы зубов , оставленными молодым тиранозавром. | Дромеозаврид |
 |
||
| Стенонихозавр | S. inequalis | Почти полный скелет и другие частичные скелеты, типовой образец | Троодонтид когда-то считался разновидностью троодона |
 |
||
Тираннозавры
| Тиранозавры из формирования парка динозавров | ||||||
|---|---|---|---|---|---|---|
| Род | Разновидность | Место нахождения | Стратиграфическое положение | Материал | Примечания | Изображений |
| Дасплетозавр | Безымянный вид | Средний и Верхний, 75.6-75 млн. Назад | Несколько экземпляров | Tyrannosaurine tyrannosaurid , также присутствует в формировании BEARPAW |

|
|
| Горгозавр | G. libratus | Нижнее-Среднее, 76,6–75,1 млн лет назад | Многочисленные экземпляры, типовой образец | Albertosaurine tyrannosaurid, также присутствует в формировании реки Джудит и , возможно образование двух медицины . |
 |
|
Цветовой ключ
|
Примечания Неопределенные или предварительные таксоны выделены мелким шрифтом ; |
Прочие рептилии
Хористодерес
Choristoderes или champsosaurs , были водные рептилии. Маленькие экземпляры были похожи на ящериц, в то время как более крупные были внешне похожи на крокодилов. В свите обнаружены остатки следующих хористодеров:
- Шампсозавр (минимум 3 вида)
- Cteniogenys
Крокодилы
В формации были обнаружены останки следующих крокодилов:
- Альберточампа
- Leidyosuchus
- как минимум 1 безымянный таксон
Ящерицы
В свите обнаружены останки следующих ящериц:
Плезиозавры
В формации были обнаружены останки следующих плезиозавров:
- Fluvionectes
- индетерминантные поликотилиды (с более короткой шеей)
Птерозавры
В свите обнаружены останки следующих птерозавров:
- Криодракон (известен по мелким и крупным экземплярам)
- 1 безымянный птерозавр, не являющийся аждархидом
Черепахи
В составе формации найдены останки следующих черепах:
- Adocus
- " Апалон "
- Aspideretoides (3 вида)
- Basilemys
- Boremys
- Judithemys
- Neurankylus
- Плезиобаена
- 2 неопределенных таксона
Млекопитающие
В свите обнаружены останки следующих млекопитающих:
-
Мультитуберкулята
- Cimexomys sp.
- Cimolodon spp.
- Cimolomys clarki
- Meniscoessus major
- Mesodma primaeva
- безымянный мультитуберкулез
-
Метатерии
- Alphadon halleyi
- Эодельфис брауни
- Э. катлери
- 5 видов « Педиомиса »
- Turgidodon russelli
- T. praesagus
-
Евтерийцы
- Cimolestes sp. (неопределенная таксономия)
- Гипсониктопс левизи
- Paranyctoides sternbergi
- Неизвестные терианы : минимум 1 вид
Рыба
В свите обнаружены остатки следующих рыб:
- Хондрихтианцы
-
Acipenseriformes ( осетровые )
- " Acipenser albertensis "
- Anchiacipenser acanthaspis
- безымянный осетр
- безымянный веслонос
-
Голосовая рыба
- Lepisosteus occidentalis ( гар )
- безымянный боуфин
- как минимум 2 других голостея
-
Костистые рыбы
- Belonostomus longirostris
- Cretophareodus ( остеоглоссоморф )
- Coriops amnicolus
- Estesesox foxi
- Oldmanesox
- Паральбула (включая Phyllodus )
- Paratarpon apogerontus ( элопоморф , как тарпон )
- по крайней мере 8 других костистых
Беспозвоночные
В свите обнаружены остатки следующих беспозвоночных:
- Пресноводные двустворчатые моллюски
- Пресноводные брюхоногие моллюски
- Campeloma (2 вида)
- Элимия
- Гониобазис (3 вида)
- Гидробия
- Lioplacodes (2 вида)
Флора
Окаменелости тела растений
В формации были обнаружены следующие окаменелости растительных тел:
- различные папоротники
- Хвойный ( Equisetaceae )
-
Голосеменные
- Платиспироксилон ( Cupressaceae )
- Подокарпоксилон ( Podocarpaceae )
- Элатоклад ( Taxodiaceae )
- Секвойя (Taxodiaceae)
- Секвойаксилон (Taxodiaceae)
- Таксодиоксилон (Taxodiaceae)
- Гинкго
-
Покрытосеменные
- Артокарпус ( Moraceae )
- Церсидифиллум ( Cercidiphyllaceae )
- Домбеопсис ( Sterculiaceae )
- Мениспермиты ( Menispermaceae )
- Пистия ( Araceae )
- Платан ( Platanaceae )
- Витис ( Vitaceae )
- Trapa ( Trapaceae )
Палиноморфы
Палиноморфы - это микрофоссилии с органическими стенками , такие как споры , пыльца и водоросли . В свите обнаружены следующие палиноморфы:
- Неизвестные производители
- не менее 8 видов
-
Chlorophyta ( зеленые водоросли и сине-зеленые водоросли )
- не менее 12 видов
-
Pyrrhophyta ( динофлагелляты , вид морских водорослей )
- неназначенные кисты
-
Мохообразные ( мхи , печеночники и роголистники )
-
Anthocerotophyta (роголистники)
- минимум 5 видов
-
Marchantiophyta (печеночники)
- не менее 14 видов
-
Bryophyta (мхи)
- минимум 5 видов
-
Anthocerotophyta (роголистники)
-
Lycopodiophyta
-
Lycopodiaceae ( клубные мхи )
- не менее 11 видов
-
Selaginellaceae (небольшие клубные мхи)
- не менее 6 видов
-
Isoetaceae ( иголки )
- минимум 1 вид
-
Lycopodiaceae ( клубные мхи )
-
Полиподиофита
-
Osmundaceae ( папоротники корицы )
- не менее 6 видов
-
Schizaeaceae ( плетистые папоротники )
- не менее 20 видов
-
Gleicheniaceae ( Gleichenia и союзники; коралловые папоротники )
- минимум 5 видов
-
Cyatheaceae ( Cyathea и союзники)
- минимум 4 вида
-
Dicksoniaceae ( Диксония и союзники)
- минимум 3 вида
-
Polypodiaceae ( папоротники )
- минимум 4 вида
-
Matoniaceae
- минимум 1 вид
-
Марсиловые
- минимум 1 вид
-
Osmundaceae ( папоротники корицы )
-
Пинофита ( голосеменные )
-
Cycadaceae ( саговники )
- минимум 3 вида
-
Caytoniaceae
- минимум 1 вид
-
Pinaceae ( сосны )
- минимум 4 вида
-
Cupressaceae ( кипарисы )
- минимум 3 вида
-
Podocarpaceae ( Podocarpus и союзники)
- минимум 4 вида
-
Хейролепидиевые
- минимум 2 вида
-
Ephedraceae ( мормонские чаи )
- не менее 6 видов
- Неизвестные голосеменные: не менее 3 видов.
-
Cycadaceae ( саговники )
-
Magnoliophyta ( покрытосеменные )
-
Магнолиопсиды ( двудольные )
-
Buxaceae ( самшит )
- минимум 1 вид
-
Гуннеровые ( gunneras )
- минимум 1 вид
-
Salicaceae ( ива , тополь , осина кряхлая )
- минимум 1 вид
-
Droseraceae ( росянки )
- минимум 1 вид
-
Olacaceae ( талловое дерево )
- минимум 2 вида
-
Loranthaceae ( эффектные омелы )
- минимум 1 вид
-
Sapindaceae ( мыльница )
- минимум 1 вид
-
Aceraceae ( клены )
- минимум 1 вид
-
Proteaceae ( протей )
- не менее 9 видов
-
Сложноцветные ( подсолнухи )
- минимум 1 вид
-
Fagaceae ( бук , дуб , каштаны )
- минимум 2 вида
-
Betulaceae ( береза , ольха )
- минимум 1 вид
-
Ulmaceae ( вязы )
- минимум 1 вид
-
Chenopodiaceae ( гусиные лапки )
- минимум 1 вид
-
Buxaceae ( самшит )
-
Лилиопсида ( однодольные )
-
Лилии ( лилии )
- не менее 6 видов
-
Cyperaceae (осоки)
- минимум 1 вид
-
Sparganiaceae ( тростник )
- возможно 1 вид
- Неизвестные покрытосеменные: не менее 88 видов.
-
Лилии ( лилии )
-
Магнолиопсиды ( двудольные )
Хронология появления новых таксонов
На следующей временной шкале показаны действительные таксоны, впервые обнаруженные у динозавра. Некоторые виды могли быть отнесены к другим родам после их первоначального описания.
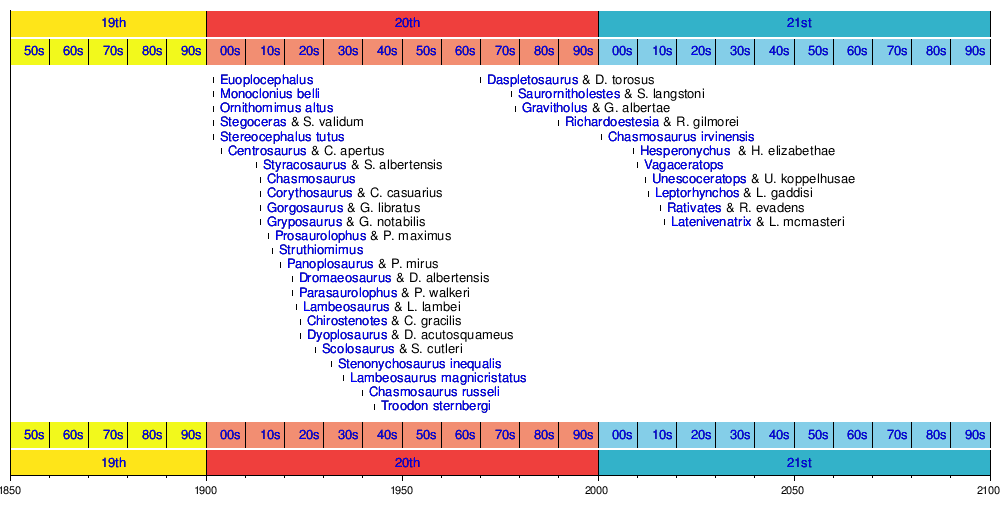
Смотрите также
Сноски
использованная литература
- Беседка, ВМ; Бернс, Мэн; Сиссонс, Р.Л. (2009). «Переописание анкилозавридного динозавра Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) и пересмотр рода». Журнал палеонтологии позвоночных . 29 (4): 1117–1135. DOI : 10.1671 / 039.029.0405 . S2CID 85665879 .
- Браман Д.Р., Коппельхус Е.Б. 2005. Кампанские палиноморфы. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 101–130.
- Бринкман, Д. Б. 2005. Черепахи: разнообразие, палеоэкология и распространение. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 202-220.
- Колдуэлл, М.В. Чешуйчатые: происхождение, филогения и палеоэкология. В: Currie, PJ, and Koppelhus, EB (eds). 2005 г. '' Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 235–248.
- Карри, П.Дж. 2005. Тероподы, в том числе птицы. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 367–397.
- Карри, П.Дж., и Коппельхус, Э.Б. (ред.). 2005. Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 648 стр.
- Эберт, Д.А. 2005. Геология. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 54–82.
- Fox, RC 2005. Млекопитающие позднего мела. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 417-435.
- К. Гао и Бринкман, Д. Б. 2005. Хористодеры из парка и его окрестностей. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 221-234.
- Гарднер, Дж. Д. 2005. Лиссамфибии. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 186-201.
- Годфри, SJ, и Карри, PJ 2005. Птерозавры. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 292-311.
- Джонстон, Пенсильвания, и Хенди, AJW, 2005. Палеоэкология моллюсков из группы верхнего мела реки Белли. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 139–166.
- Коппельхус, Е.Б. 2005. Палеоботаника. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 131–138.
- Лексикон канадских геологических единиц. «Формирование парка динозавров» . Архивировано из оригинала на 2013-02-21 . Проверено 29 марта 2011 .
- Нойман, А.Г., Бринкман, Д.Б. 2005. Рыбы речных пластов. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 167–185.
- Райан, MJ, и Эванс, округ Колумбия, 2005. Орнитисхийские динозавры. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 312-348.
- Сато Т., Эберт Д.А., Николлс Е.Л. и Манабе М. 2005. Остатки плезиозавров от неморских до паралических отложений. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 249-276.
- Танке, Д.Х. и Бретт-Сурман, М.К. 2001. Свидетельства появления птенцов и гадрозавров размером с птенцов (Reptilia: Ornithischia) из провинциального парка динозавров (формирование парка динозавров: кампания), Альберта, Канада. С. 206–218. В: Мезозойская жизнь позвоночных - новые исследования, вдохновленные палеонтологией Филипа Дж. Карри. Под редакцией Д.Х. Танке и К. Карпентера. Издательство Индианского университета: Блумингтон. xviii + 577 с.
- Сяо-Чун Ву. 2005. Крокодилы. В: Карри, П.Дж., и Коппельхус, Б.Б. (редакторы), Провинциальный парк динозавров: обнаружена впечатляющая древняя экосистема. Издательство Индианского университета: Блумингтон и Индианаполис, 277-291