
Рыбий плавник -Fish fin
.png)
(1) грудные плавники (парные), (2) брюшные плавники (парные), (3) спинной плавник ,
(4) жировой плавник, (5) анальный плавник, (6) хвостовой ( хвост) плавник
Плавники представляют собой отличительные анатомические особенности, состоящие из костных шипов или лучей , выступающих из тела рыбы . Они покрыты кожей и соединены вместе либо перепончатым образом, как у большинства костистых рыб , либо похожим на плавник , как у акул . Кроме хвостового или хвостового плавника , плавники рыб не имеют прямой связи с позвоночником и поддерживаются только мышцами . Их основная функция заключается в том, чтобы помочь рыбе плавать .
Плавники, расположенные в разных местах на рыбе, служат разным целям, таким как движение вперед, поворот, удержание вертикального положения или остановка. Большинство рыб используют плавники при плавании, летучие рыбы используют грудные плавники для планирования, а лягушки используют их для ползания. Плавники можно использовать и для других целей; самцы акул и рыб- москитов используют модифицированный плавник для выделения спермы, акулы-молотильщики используют свой хвостовой плавник, чтобы оглушить добычу, у рифовых каменных рыб есть шипы в спинных плавниках, которые впрыскивают яд, удильщики используют первый шип своего спинного плавника как удочку, чтобы заманить добычу. , а спинороги избегают хищников , протискиваясь в коралловые расщелины и используя шипы в своих плавниках, чтобы зафиксироваться на месте.
Типы плавников
Для каждого типа плавников существует ряд видов рыб, у которых этот конкретный плавник был утерян в процессе эволюции.
| Грудные плавники |

|
Парные грудные плавники расположены по бокам, обычно сложены сразу за жаберной крышкой и гомологичны передним конечностям четвероногих .
|
|---|---|---|
|
Брюшные плавники (брюшные плавники) |

|
Парные тазовые или брюшные плавники обычно располагаются вентрально ниже и позади грудных плавников, хотя у многих семейств рыб они могут располагаться перед грудными плавниками (например, у трески). Они гомологичны задним конечностям четвероногих . Брюшной плавник помогает рыбе двигаться вверх или вниз по воде, резко поворачиваться и быстро останавливаться.
|
| Спинной плавник |
 |

Спинной плавник голавля ( Leuciscus cephalus )
Спинные плавники расположены на спине. У рыбы может быть до трех спинных плавников. Спинные плавники служат для защиты рыбы от перекатывания и помогают ей при резких поворотах и остановках.
|
| Анальный/клоакальный плавник |

|
Анальный /клоакальный плавник расположен на брюшной поверхности позади ануса / клоаки . Этот плавник используется для стабилизации рыбы во время плавания. |
| Жировой плавник |
 |
Жировой плавник представляет собой мягкий мясистый плавник, расположенный на спине позади спинного плавника и прямо перед хвостовым плавником. Он отсутствует у многих семейств рыб, но встречается у девяти из 31 отряда эвтелеостевых ( Percopsiformes , Myctophiformes , Aulopiformes , Stomiiformes , Salmoniformes , Osmeriformes , Characiformes , Siluriformes и Argentiniformes ). Известными представителями этих отрядов являются лососевые , харациновые и сомы .
Функция жирового плавника остается загадкой. Его часто обрезают, чтобы отметить рыбу, выращенную в заводских условиях, хотя данные за 2005 год показали, что у форели с удаленным жировым плавником частота ударов хвостом на 8% выше. Дополнительная информация, опубликованная в 2011 году, предполагает, что плавник может иметь жизненно важное значение для обнаружения и реагирования на такие раздражители, как прикосновение, звук и изменения давления. Канадские исследователи обнаружили в плавнике нейронную сеть, что указывает на то, что она, вероятно, имеет сенсорную функцию, но до сих пор не уверены, каковы будут последствия ее удаления. Сравнительное исследование, проведенное в 2013 году, показывает, что жировой плавник может развиваться двумя разными способами. Один из них — лососевидный, при котором жировой плавник развивается из личиночно-плавниковой складки одновременно и таким же непосредственным образом, как и остальные срединные плавники. Другой - харациформный путь, при котором жировой плавник развивается поздно после того, как уменьшилась личиночно-плавниковая складка и развились другие срединные плавники. Они утверждают, что существование харациформного типа развития предполагает, что жировой плавник - это не «просто остаток складки личиночного плавника», и это несовместимо с мнением о том, что жировой плавник не имеет функции. Исследования, опубликованные в 2014 году, показывают, что жировой плавник неоднократно развивался в отдельных линиях . |
| Хвостовой плавник (хвостовой плавник) |
 
|
Хвостовой плавник — это хвостовой плавник (от латинского cauda — хвост), расположенный на конце хвостового стебля и служащий для движения. См. движение тела и хвостового плавника .
(A) - Гетероцеркальный означает, что позвонки доходят до верхней доли хвоста, делая его длиннее (как у акул ). Противоположно гипоцеркальному.
(B) - Протоцеркальный означает, что позвонки доходят до кончика хвоста, а хвост симметричен, но не расширен (как у первых рыб и круглоротых , а также у более примитивного предшественника у ланцетников ) . (C) - Гомоцеркальный , когда плавник кажется внешне симметричным, но на самом деле позвонки простираются на очень короткое расстояние в верхнюю долю плавника. (D) - Дифицеркальный означает, что позвонки доходят до кончика хвоста, а хвост симметричный и расширенный (как у бичира , двоякодышащей рыбы , миноги и латимерии ). Большинство палеозойских рыб имели дифицеркальный гетероцеркальный хвост. У большинства современных рыб ( костистых ) хвост гомоцеркальный. Они появляются в различных формах и могут появляться:
|
| Хвостовой киль |
_diagram_cropped.GIF)
|
Некоторые виды быстроплавающих рыб имеют горизонтальный хвостовой киль прямо перед хвостовым плавником. Подобно килю корабля, это боковой гребень на хвостовом стебле, обычно состоящий из щитков (см. Ниже), который обеспечивает устойчивость и поддержку хвостового плавника. Киль может быть один парный, по одному с каждой стороны, или две пары сверху и снизу.
Плавники - это маленькие плавники, обычно позади спинного и анального плавников (у бихировых есть только плавники на спинной поверхности и нет спинного плавника). У некоторых рыб, таких как тунец или сайра , они не имеют лучей, не убираются и находятся между последним спинным и/или анальным плавником и хвостовым плавником. |
Костистые рыбы

Костистые рыбы образуют таксономическую группу под названием Osteichthyes. Их скелет состоит из кости, и их можно сравнить с хрящевыми рыбами , скелеты которых состоят из хрящей . Костные рыбы делятся на лучепёрых и кистеперых . Большинство рыб лучеперые, чрезвычайно разнообразная и многочисленная группа, состоящая из более чем 30 000 видов. На сегодняшний день это самый большой класс позвоночных. В далеком прошлом кистепёрые рыбы были в изобилии. В настоящее время они в основном вымерли, осталось только восемь живых видов. У костистых рыб есть шипы на плавниках и лучи, называемые лепидотрихиями. У них обычно есть плавательный пузырь , который позволяет рыбе создавать нейтральный баланс между погружением и плаванием, не используя плавники. Однако плавательные пузыри отсутствуют у многих рыб, в первую очередь у двоякодышащих рыб , которые являются единственными рыбами, сохранившими примитивные легкие, присутствующие у общего предка костистых рыб, от которых развился плавательный пузырь. У костистых рыб также есть жаберная крышка , которая помогает им дышать, не используя плавники для плавания.
лопастные плавники

Рыбы с кистеперыми плавниками образуют класс костных рыб , называемых саркоптеригиями. У них мясистые, лопастные , парные плавники, которые соединены с телом одной костью. Плавники лопастных рыб отличаются от плавников всех других рыб тем, что каждый из них держится на мясистом, лопастном, чешуйчатом стебле, отходящем от тела. Грудные и брюшные плавники имеют сочленения, напоминающие конечности четвероногих. Эти плавники превратились в ноги первых четвероногих наземных позвоночных, земноводных . У них также есть два спинных плавника с отдельными основаниями, в отличие от одного спинного плавника лучепёрых рыб .
Латимерия — рыба с лопастными плавниками, сохранившаяся до наших дней. Считается, что примерно 408 миллионов лет назад, в раннем девоне, он эволюционировал в свою нынешнюю форму. Передвижение латимерий уникально для их вида. Чтобы передвигаться, латимерии чаще всего используют восходящие или нисходящие потоки и дрейф. Они используют свои парные плавники, чтобы стабилизировать свое движение в воде. На дне океана их парные плавники не используются ни для каких движений. Целаканты могут создавать тягу для быстрого старта, используя свои хвостовые плавники. Благодаря большому количеству плавников латимерии обладают высокой маневренностью и могут ориентировать свое тело практически в любом направлении в воде. Было замечено, как они делают стойку на голове и плавают животом вверх. Считается, что их ростральный орган помогает латимериям получать электровосприятие, которое помогает им двигаться вокруг препятствий.
Двоякодышащие рыбы также являются живыми кистеперыми рыбами. Встречаются в Африке ( Protopterus ), Австралии ( Neoceratodus ) и Южной Америке ( Lepidosiren ).
Разнообразие плавников у кистепёрых рыб

Пятнистая двоякодышащая рыба Protopterus dolloi
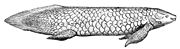
Квинслендская двоякодышащая рыба Neoceratodus forsteri





Лучевые плавники

Рыбы с лучистыми плавниками образуют класс костных рыб , называемых Actinopterygii. Их плавники содержат шипы или лучи. Плавник может содержать только колючие лучи, только мягкие лучи или их комбинацию. Если присутствуют оба, колючие лучи всегда впереди . Шипы обычно жесткие и острые. Лучи обычно мягкие, гибкие, сегментированные и могут быть разветвленными. Эта сегментация лучей является основным отличием, отделяющим их от шипов; шипы могут быть гибкими у некоторых видов, но они никогда не будут сегментированы.
Шипы имеют множество применений. У сома они используются как форма защиты; у многих сомов есть способность запирать шипы наружу. Спинороги также используют шипы, чтобы запираться в щелях, чтобы их не вытащили.
Lepidotrichia обычно состоят из кости , но у ранних остейхтианов , таких как Cheirolepis , были также дентин и эмаль . Они сегментированы и выглядят как ряд дисков, уложенных друг на друга. Они могли быть получены из кожных чешуек. Считается, что генетической основой формирования плавниковых лучей являются гены, кодирующие выработку определенных белков. Было высказано предположение, что эволюция конечностей четвероногих от лопастных рыб связана с потерей этих белков.
Разнообразие плавников у лучепёрых рыб
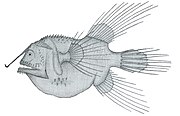
Удильщик-фанфин Caulophryne jordani

Стройная солнечная рыба Ranzania laevis

Веера Pteraclis carolinus

Прозрачный топорик Sternoptyx diaphana

Серебряный грубый Hoplostethus mediterraneus

Рыба-нож Equetus lanceolatus

Атлантический бриз Брама Брама

Атлантический затонувший рыба Polyprion americanus

Звездчатая рыба -фугу Arothron stellatus
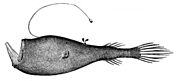
Созерцающий звезды морской черт Ceratias uranoscopus

Риджхед Поромитра единорогис

Тропическая двукрылая летучая рыба Exocoetus evolans

Кусковый угорь Benthocometes robustus

Крысохвост Trachonurus sulcatus

Рыба- тренога Bathypterois grallator

Гигантская гребенчатая рыба Regalecus glesne

Короткоклювая подводная рыба Tetrapturus angustirostris

Рыба-нож- призрак Sternarchorhynchus oxyrhynchus

Ремора Remora brachyptera

Горный шкипер с голубыми пунктирами Blenniella periophthalmus

Нильский бичир Polypterus bichir

Прибрежная головорезная форель Oncorhynchus clarkii

Африканский масляный сом Schilbe mystus


.gif)














.gif)
.gif)




Хрящевые рыбы


Хрящевые рыбы образуют класс рыб под названием Chondrichthyes. Их скелеты состоят из хрящей , а не костей . В класс входят акулы , скаты и химеры . Скелеты акульих плавников удлинены и поддерживаются мягкими и несегментированными лучами, называемыми цератотрихиями, нитями эластичного белка, напоминающими роговой кератин в волосах и перьях. Первоначально грудной и тазовый пояса, не содержащие кожных элементов, не соединялись. У более поздних форм каждая пара плавников стала вентрально соединенной посередине, когда развились лопаточно-коракоидные и лобково-шадные перемычки. У скатов грудные плавники соединены с головой и очень гибкие. Одной из основных характеристик, присутствующих у большинства акул, является гетероцеркальный хвост, который помогает в передвижении. У большинства акул восемь плавников. Акулы могут дрейфовать только от объектов прямо перед ними, потому что их плавники не позволяют им двигаться хвостом вперед.
Как и у большинства рыб, хвосты акул создают толчок, поэтому скорость и ускорение зависят от формы хвоста. Форма хвостового плавника значительно различается у разных видов акул из-за их эволюции в разных средах. Акулы обладают гетероцеркальным хвостовым плавником, у которого спинная часть обычно заметно больше брюшной . Это связано с тем, что позвоночник акулы простирается в эту спинную часть, обеспечивая большую площадь поверхности для прикрепления мышц . Это позволяет более эффективно передвигаться среди этих хрящевых рыб с отрицательной плавучестью . Напротив, у большинства костистых рыб хвостовой плавник гомоцеркальный .
Тигровые акулы имеют большую верхнюю долю , что позволяет плавать медленно и резко увеличивать скорость. Тигровая акула должна легко извиваться и поворачиваться в воде во время охоты, чтобы поддерживать свою разнообразную диету, в то время как акула-порбигль , которая охотится на стайную рыбу, такую как скумбрия и сельдь , имеет большую нижнюю долю, которая помогает ей не отставать от своей быстрой рыбы. - плавающая добыча. Другие приспособления хвоста помогают акулам более точно ловить добычу, например, акула-молотилка использует свою мощную удлиненную верхнюю долю для оглушения рыбы и кальмаров.
Акульи плавники
По данным Humane Society International , около 100 миллионов акул убивают каждый год из-за их плавников, что известно как акульи плавники . После отрезания плавников изуродованных акул бросают обратно в воду и оставляют умирать.
В некоторых странах Азии акульи плавники являются кулинарным деликатесом, например супом из акульих плавников . В настоящее время международные опасения по поводу устойчивости и благополучия акул повлияли на потребление и доступность супа из акульих плавников во всем мире. Вылов акульих плавников запрещен во многих странах.
Функции ребра
Создание тяги
Плавники в форме фольги создают тягу при движении, подъем плавника приводит в движение воду или воздух и толкает плавник в противоположном направлении. Водные животные получают значительную тягу , двигая плавниками взад и вперед в воде. Часто используется хвостовой плавник , но некоторые водные животные создают толчок грудными плавниками .

Кавитация возникает, когда отрицательное давление вызывает образование пузырьков (полостей) в жидкости, которые затем быстро и сильно схлопываются. Это может привести к значительному повреждению и износу. Кавитационное повреждение может произойти с хвостовыми плавниками мощных плавающих морских животных, таких как дельфины и тунец. Кавитация чаще всего возникает у поверхности океана, где давление окружающей воды относительно низкое. Даже если у них есть способность плавать быстрее, дельфинам, возможно, придется ограничить свою скорость, потому что схлопывающиеся кавитационные пузыри на их хвосте слишком болезненны. Кавитация также замедляет тунца, но по другой причине. В отличие от дельфинов, эти рыбы не чувствуют пузырьков, потому что у них костные плавники без нервных окончаний. Тем не менее, они не могут плыть быстрее, потому что кавитационные пузырьки создают паровую пленку вокруг их плавников, ограничивающую их скорость. На тунце были обнаружены повреждения, соответствующие кавитационному повреждению.
Скумбридовые рыбы (тунец, скумбрия и бонито) особенно хорошо плавают. Вдоль края в задней части их тела проходит линия маленьких неубирающихся плавников без лучей, известных как плавники . Было много предположений о функции этих плавников. Исследования, проведенные в 2000 и 2001 годах Науэном и Лаудером, показали, что «плавники оказывают гидродинамическое влияние на локальный поток во время устойчивого плавания» и что «самый задний плавник ориентирован на перенаправление потока в развивающийся хвостовой вихрь, что может увеличить тягу, создаваемую плавником. хвост плавающей скумбрии».
Рыбы используют несколько плавников, поэтому возможно, что данный плавник может иметь гидродинамическое взаимодействие с другим плавником. В частности, плавники непосредственно перед хвостовым (хвостовым) плавником могут быть ближайшими плавниками, которые могут напрямую влиять на динамику потока в хвостовом плавнике. В 2011 году исследователи, использующие методы объемного изображения , смогли создать «первые мгновенные трехмерные изображения структур следа, создаваемых свободно плавающими рыбами». Они обнаружили, что «непрерывные удары хвоста привели к образованию связанной цепочки вихревых колец» и что «следы спинного и анального плавников быстро увлекаются следом хвостового плавника, примерно в пределах временных рамок последующего удара хвостом».
Управление движением
Как только движение установлено, само движение можно контролировать с помощью других плавников.


.jpg)
Форма тела рифовых рыб часто отличается от формы тела рыб в открытой воде . Рыбы в открытой воде обычно созданы для скорости, они имеют обтекаемую форму, как торпеды, чтобы свести к минимуму трение при движении в воде. Рифовые рыбы действуют в относительно замкнутых пространствах и сложных подводных ландшафтах коралловых рифов . Для этого маневренность важнее скорости по прямой, поэтому коралловые рифовые рыбы имеют развитые тела, которые оптимизируют их способность бросаться и менять направление. Они перехитрили хищников, уворачиваясь от трещин в рифе или играя в прятки вокруг голов кораллов. Грудные и тазовые плавники многих рифовых рыб, таких как рыба - бабочка , рыба- ласточка и рыба- ангел , эволюционировали таким образом, что они могут действовать как тормоза и позволяют выполнять сложные маневры. Многие рифовые рыбы, такие как рыба - бабочка , рыба- ласточка и рыба- ангел , развили тела, которые являются глубокими и сжатыми с боков, как блины, и могут входить в трещины в скалах. Их тазовые и грудные плавники эволюционировали по-разному, поэтому они действуют вместе со сплющенным телом, чтобы оптимизировать маневренность. Некоторые рыбы, такие как рыба-фугу, рыба - филе и рыба- хобот , полагаются на грудные плавники для плавания и почти не используют хвостовые плавники.
Воспроизведение


Самцы хрящевых рыб (акулы и скаты), а также самцы некоторых живородящих лучеперых рыб имеют плавники, которые были изменены, чтобы функционировать как внутренние органы , репродуктивные придатки, обеспечивающие внутреннее оплодотворение . У лучепёрых рыб они называются гоноподиями или андроподиями , а у хрящевых — класперами .
Гоноподии встречаются у самцов некоторых видов семейств Anablepidae и Poeciliidae . Это анальные плавники, которые были модифицированы для работы в качестве подвижных внутренних органов и используются для оплодотворения самок молоком во время спаривания. Третий, четвертый и пятый лучи анального плавника самца образуют трубчатую структуру, в которую выбрасывается сперма рыб. Когда гоноподий готов к спариванию, он становится прямостоячим и направлен вперед в сторону самки. Самец вскоре вставляет орган в половое отверстие самки с крючкообразными приспособлениями, которые позволяют рыбе цепляться за самку, чтобы обеспечить оплодотворение. Если самка остается неподвижной, а ее партнер касается ее анального отверстия своим гоноподием, она оплодотворяется. Сперма сохраняется в яйцеводе самки. Это позволяет самкам оплодотворять себя в любое время без дополнительной помощи со стороны самцов. У некоторых видов гоноподий может составлять половину общей длины тела. Иногда плавник слишком длинный, чтобы его можно было использовать, как у «лирохвостых» пород Xiphophorus helleri . У самок, получавших лечение гормонами, могут развиться гоноподии. Они бесполезны для разведения.
Подобные органы с аналогичными характеристиками встречаются и у других рыб, например, андроподий у Hemirhamphodon или у Goodeidae или гоноподий в среднем триасе † Saurichthys , самый старый известный пример живорождения у лучепёрых рыб.
Кламперы встречаются на самцах хрящевых рыб . Они представляют собой заднюю часть брюшных плавников, которые также были модифицированы, чтобы функционировать как внутренние органы, и используются для направления спермы в клоаку самки во время совокупления. Акт спаривания у акул обычно включает поднятие одной из застежек, чтобы вода попала в сифон через определенное отверстие . Затем застежка вставляется в клоаку, где она открывается, как зонтик, чтобы зафиксировать свое положение. Затем сифон начинает сокращаться, выталкивая воду и сперму.
Другие функции
Другие виды использования плавников включают ходьбу и сидение на морском дне, скольжение по воде, охлаждение температуры тела, оглушение добычи, демонстрацию (отпугивание хищников, ухаживание), защиту (ядовитые шипы плавников, зацепление между кораллами), заманивание добычи, и структур прикрепления.
Индо - тихоокеанский парусник имеет выдающийся спинной плавник. Подобно скумбриям и другим морским рыбам , они обтекаются, втягивая спинные плавники в бороздку на теле, когда плывут . Огромный спинной плавник, или парус, парусника большую часть времени держится втянутым. Парусники поднимают их, если хотят загнать косяк мелкой рыбы, а также после периодов высокой активности, предположительно, чтобы остыть.
_in_Waikiki_Aquarium.JPG)




У восточного морского морского петуха большие грудные плавники, которые он обычно прижимает к телу и расширяется, когда ему угрожают, чтобы напугать хищников. Несмотря на свое название, это демерсальная рыба , а не летающая рыба, и она использует свои брюшные плавники, чтобы ходить по дну океана.
Плавники могут иметь приспособительное значение как половые украшения. Во время ухаживания самка цихлиды , Pelvicachromis taeniatus , демонстрирует большой и привлекающий внимание фиолетовый брюшной плавник . «Исследователи обнаружили, что самцы явно предпочитали самок с большим брюшным плавником, и что брюшные плавники вырастали более непропорционально, чем другие плавники у самок рыб».






Эволюция
Эволюция парных плавников
Есть две преобладающие гипотезы, которые исторически обсуждались как модели эволюции парных плавников у рыб: теория жаберных дуг и теория боковых плавниковых складок. Первая, обычно называемая « гипотезой Гегенбаура », была выдвинута в 1870 году и предполагает, что «парные плавники произошли от жаберных структур». Это потеряло популярность в пользу теории боковых плавниковых складок, впервые предложенной в 1877 году, которая предполагает, что парные плавники отпочковались от продольных боковых складок эпидермиса сразу за жабрами. Обе гипотезы слабо подтверждаются летописью окаменелостей и эмбриологией. Однако недавние открытия в области формирования паттернов развития побудили пересмотреть обе теории, чтобы лучше понять происхождение парных плавников.
Классические теории
Концепция «архиптеригиума» Карла Гегенбаура была введена в 1876 году. Он описывался как жаберный луч или «соединенный хрящевой стебель», отходящий от жаберной дуги. Дополнительные лучи отходили вдоль дуги и от центрального жаберного луча. Гегенбаур предложил модель трансформационной гомологии , согласно которой все парные плавники и конечности позвоночных были трансформациями Archipterygium. Основываясь на этой теории, парные придатки, такие как грудные и брюшные плавники, должны были дифференцироваться от жаберных дуг и мигрировать назад. Однако в летописи окаменелостей эта гипотеза получила ограниченное подтверждение как морфологически, так и филогенетически. Кроме того, почти не было свидетельств передне-задней миграции брюшных плавников. Такие недостатки теории жаберных дуг привели к ее ранней кончине в пользу теории боковой плавниковой складки, предложенной Сент-Джорджем Джексоном Мивартом , Фрэнсисом Бальфуром и Джеймсом Кингсли Тэчером .
Теория боковой плавниковой складки предполагала, что парные плавники развились из боковых складок вдоль стенки тела рыбы. Точно так же, как сегментация и отпочкование срединной плавниковой складки дали начало срединным плавникам, было высказано предположение, что аналогичный механизм сегментации и удлинения плавниковых зачатков от латеральной плавниковой складки привел к парным грудным и брюшным плавникам. Однако в летописи окаменелостей было мало свидетельств перехода от боковой складки к плавнику. Кроме того, позже было филогенетически продемонстрировано, что грудные и брюшные плавники имеют разное эволюционное и механистическое происхождение.
Эволюционная биология развития
Недавние исследования онтогенеза и эволюции парных придатков сравнивали безплавниковых позвоночных, таких как миноги , с хондрихиями , самыми базальными из ныне живущих позвоночных с парными плавниками. В 2006 году исследователи обнаружили, что такое же генетическое программирование, связанное с сегментацией и развитием срединных плавников, было обнаружено в развитии парных придатков у кошачьих акул . Хотя эти находки напрямую не подтверждают гипотезу боковой складки плавников, первоначальная концепция эволюционного механизма общего парного срединного плавника остается актуальной.
Аналогичное обновление старой теории можно найти в программировании развития жаберных дуг и парных придатков хондрихий. В 2009 году исследователи из Чикагского университета продемонстрировали, что существуют общие механизмы формирования молекулярных паттернов на раннем этапе развития жаберной дуги и парных плавников. Находки, подобные этим, побудили пересмотреть некогда опровергнутую теорию жаберных дуг.
От плавников к конечностям
Рыбы являются предками всех млекопитающих, рептилий, птиц и амфибий. В частности, наземные четвероногие (четвероногие животные) произошли от рыб и совершили свои первые набеги на сушу 400 миллионов лет назад. Для передвижения они использовали парные грудные и брюшные плавники. Грудные плавники превратились в передние ноги (руки в случае людей), а брюшные плавники превратились в задние ноги. Большая часть генетического механизма, который строит ходячую конечность у четвероногих, уже присутствует в плавниках рыб.
Аристотель признал различие между аналогичными и гомологичными структурами и сделал следующее пророческое сравнение: «Птицы чем-то напоминают рыб. У птиц крылья в верхней части тела, а у рыб два плавника в передней части тела. У птиц ноги расположены на нижней части тела, а у большинства рыб есть вторая пара плавников на нижней части тела и рядом с передними плавниками».
- Аристотель, De incessu animalium


В 2011 году исследователи из Университета Монаш в Австралии использовали примитивных, но все еще живых двоякодышащих рыб , «чтобы проследить эволюцию мышц тазовых плавников и выяснить, как развивались несущие нагрузку задние конечности четвероногих». Дальнейшие исследования в Чикагском университете показали, что у двоякодышащих рыб, идущих по дну, уже развились характеристики походки наземных четвероногих.
В классическом примере конвергентной эволюции грудные конечности птерозавров , птиц и летучих мышей в дальнейшем эволюционировали по независимым путям в летающие крылья. Даже у летающих крыльев есть много общего с ходячими ногами, и основные аспекты генетической схемы грудного плавника были сохранены.
Первые млекопитающие появились в пермский период (между 298,9 и 252,17 млн лет назад). Несколько групп этих млекопитающих начали возвращаться в море, в том числе китообразные (киты, дельфины и морские свиньи). Недавний анализ ДНК показывает, что китообразные произошли от парнокопытных и что у них есть общий предок с бегемотом . Около 23 миллионов лет назад в море начала возвращаться другая группа медвежьих наземных млекопитающих. Это были тюлени . То, что стало ходячими конечностями у китообразных и тюленей, независимо эволюционировало в новые формы плавников. Передние конечности превратились в ласты , а задние были либо утрачены (китообразные), либо также преобразованы в ласты (ластоногие). У китообразных на конце хвоста появилось два плавника, называемых трематодами . Хвосты рыб обычно вертикальные и двигаются из стороны в сторону. Сосальщики китообразных расположены горизонтально и двигаются вверх и вниз, потому что иглы китообразных изгибаются так же, как и у других млекопитающих.

Ихтиозавры — древние рептилии, напоминавшие дельфинов. Впервые они появились около 245 миллионов лет назад и исчезли около 90 миллионов лет назад.
«Эта морская рептилия с наземными предками настолько сильно конвергировала с рыбами, что фактически развила спинной плавник и хвостовой плавник для улучшения передвижения в воде. Эти структуры тем более примечательны, что они произошли из ничего — у предковой наземной рептилии не было горба его спина или лезвие на хвосте, чтобы служить предшественником».
Биолог Стивен Джей Гулд сказал, что ихтиозавр был его любимым примером конвергентной эволюции .
Плавники или ласты различной формы и в разных местах (конечностях, теле, хвосте) также развились у ряда других групп четвероногих, включая ныряющих птиц, таких как пингвины (измененные из крыльев), морские черепахи (передние конечности преобразованы в ласты), мозазавры. (конечности превращены в ласты) и морские змеи (вертикально расширенный, уплощенный хвостовой плавник).
Роботизированные плавники

Использование плавников для движения водных животных может быть чрезвычайно эффективным. Было подсчитано, что некоторые рыбы могут достичь эффективности движения более 90%. Рыба может ускоряться и маневрировать намного эффективнее, чем лодки или подводные лодки , и производить меньше волнения и шума на воде. Это привело к биомиметическим исследованиям подводных роботов, которые пытаются имитировать передвижение водных животных. Примером может служить робот-тунец, созданный Институтом полевой робототехники для анализа и математического моделирования грозоподобного движения . В 2005 году в лондонском аквариуме Sea Life были представлены три рыбы-робота, созданные отделом компьютерных наук Университета Эссекса . Рыбы были разработаны, чтобы быть автономными, плавать и избегать препятствий, как настоящие рыбы. Их создатель утверждал, что пытался совместить «скорость тунца, ускорение щуки и навигационные навыки угря».
AquaPenguin , разработанный немецкой компанией Festo , копирует обтекаемую форму и движение передних ласт пингвинов . Festo также разработала AquaRay , AquaJelly и AiraCuda , имитирующие передвижение скатов манта, медуз и барракуд соответственно.
В 2004 году Хью Херр из Массачусетского технологического института создал прототип биомехатронного робота-рыбы с живым приводом , хирургическим путем пересадив роботу мышцы лягушачьих лапок, а затем заставив робота плавать, возбуждая мышечные волокна электричеством.
Роботизированные рыбы предлагают некоторые исследовательские преимущества, такие как возможность исследовать отдельные части конструкции рыбы отдельно от остальной части рыбы. Однако это может привести к чрезмерному упрощению биологии, поэтому ключевые аспекты дизайна животных будут упущены из виду. Роботизированные рыбы также позволяют исследователям изменять один параметр, такой как гибкость или конкретное управление движением. Исследователи могут напрямую измерять силы, что нелегко сделать с живой рыбой. «Роботизированные устройства также облегчают трехмерные кинематические исследования и коррелированный гидродинамический анализ, поскольку можно точно знать расположение локомоторной поверхности. Кроме того, можно запрограммировать отдельные компоненты естественного движения (например, прямой и обратный ход машущего придатка). отдельно, чего, безусловно, трудно добиться при работе с живым животным».
Смотрите также
- Головоногий плавник
- Движение плавников и ласт
- Передвижение рыбы
- Полидактилия у ранних четвероногих
- РобоТуна
- Суп из акульих плавников
- Компромиссы для передвижения в воздухе и воде
- Волнообразная локомоция
Рекомендации
Цитаты
Список используемой литературы
-
Гамлет, Уильям С. (1999). Акулы, скаты и скаты: биология пластиножаберных рыб (1-е изд.). стр. 56: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-6048-5.
{{cite book}}: CS1 maint: местоположение ( ссылка )
дальнейшее чтение
- Холл, Брайан К. (2007) Плавники превращаются в конечности: эволюция, развитие и трансформация Чикагский университет Press. ISBN 9780226313375 .
- Хелфман Г., Коллетт Б. Б., Фейси Д. Е. и Боуэн Б. В. (2009) «Функциональная морфология передвижения и питания» , глава 8, стр. 101–116. В: Разнообразие рыб: биология , John Wiley & Sons. ISBN 9781444311907 .
- Лаудер, Г.В. ; Науэн, Дж. К.; Друкер, Э. Г. (2002). «Экспериментальная гидродинамика и эволюция: функция срединных плавников у лучеперых рыб» . интегр. Комп. биол . 42 (5): 1009–1017. дои : 10.1093/icb/42.5.1009 . PMID 21680382 .
- Лаудер, Г.В.; Друкер, Э. Г. (2004). «Морфология и экспериментальная гидродинамика управляющих поверхностей плавников рыб» (PDF) . Журнал океанической инженерии . 29 (3): 556–571. Бибкод : 2004IJOE...29..556L . doi : 10.1109/joe.2004.833219 . S2CID 36207755 .
Внешние ссылки
- Гомология плавниковых лепидотрихий у остейхтиевых рыб
- Паутина земной жизни рыбьего плавника
- Могут ли роботы-рыбы находить загрязнения? Как работает . По состоянию на 30 января 2012 г.